 Antigènes
Antigènes
Le système Forssman est constitué d'un seul antigène de grande fréquence : FORS. Cet antigène a été appelé ainsi car c'est le médecin suédois John F. Forssman qui l'a découvert en 1911.
Biosynthèse
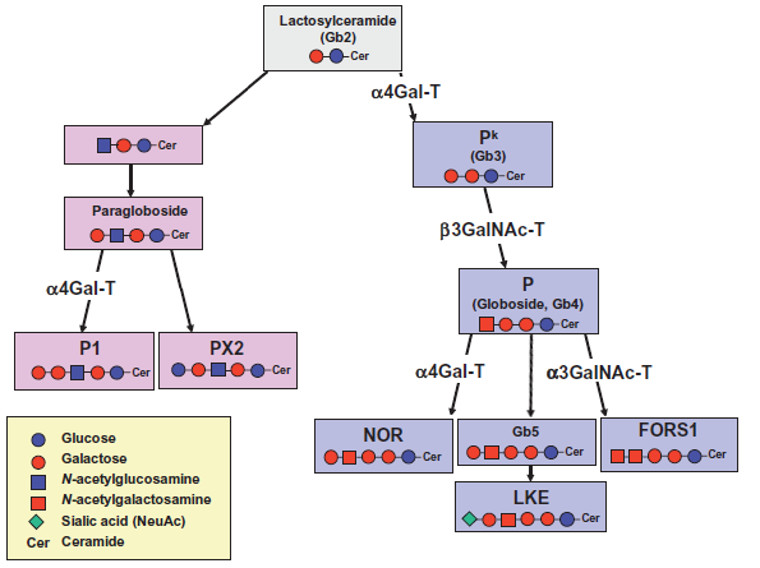
Le gène GBGT1 du chromosome 9 code pour une GBGT1 synthétase (glycosyltransferase) qui est à l'origine de l'antigène Fors. A la suite d'une série de réactions de glycosylation, l'enzyme codée par le gène catalyse le transfert du GalNAc de l'UDP-GalNAc nucléotide-sucre au résidu GalNAc terminal du globoside (GalNAcβ1-> 3Galα1-> 4Galβ1-> 4Glcβ1-> 1CER). Cet antigène se termine donc avec α-3-N-acétyl-D-galactosamine.
Antigène
Cet antigène de faible fréquence n'est pas exclusivement érythrocytaire et a été décrit dans diverses espèces animales, les plantes et les bactéries. Il est résistant aux différents traitements enzymatiques.
En 1987, 3 familles anglaises présentaient un nouvel antigène Apae. Mais ce nouveau groupe Apae vient d'être remis en cause avec l'étude du système de FORS en 2012. En effet, les patients Apae étaient de génotype O/O. Les scientifiques pensent que les patients de groupe Apae sont en fait des patients Fors positif, étiquetés Apae car les antisérum A réagissaient avec l'antigène Fors présentant la même terminaison (α-3-N-acétyl-D-galactosamine).
Le phénotype FORS1 négatif résulte d'un allèle variant conduisant à la synthèse d'une glycosyltransférase inactive.
 Anticorps
Anticorps
Les patients FORS1 négatif ont naturellement l'anticorps correspondant, stimulé par les bactéries de la flore intestinale. Ces anticorps sont généralement de classe IgM et parfois IgG. Ils présentent une réactivité optimale à 4°C. Certains d'entre eux sont détectables en test indirect à l'antiglobuline (TIA), et d'autres sont aggluninants avec une réactivité très variable d'un individu à l'autre.
Leur impact en cas de transfusions incompatibles ou de grossesses n'est pas connu. Il a été démontré que l'anti-FORS avait une capacité à hémolyser les hématies in vitro en présence de complément surtout lorsqe les hématies sont papaïnées.