L'immunité peut être définie comme l'ensemble des mécanismes biologiques permettant à un organisme de reconnaître et de tolérer ce qui lui appartient (le soi), et de reconnaître et de rejeter ce qui lui est étranger (le non soi) : les substances étrangères ou les agents infectieux auxquels il est exposé, mais aussi ses propres constituants altérés (comme des cellules tumorales).
L'immunité met en jeu deux processus étroitement imbriqués :
- l'immunité non spécifique, d'action immédiate, qui peut faire intervenir des cellules responsables de la phagocytose
- l'immunité spécifique, qui se développe en quelques jours et dépend de la reconnaissance spécifique de la substance étrangère, prélude à sa destruction ; elle garde le souvenir de la rencontre.
Immunité non spécifique
C'est l'ensemble de moyens de défense non spécifiques à un germe donné. Elle comprend les défenses de surface (peau, trachée, intestin) qui représentent la barrière du corps humain vis à vis des agents pathogènes extérieurs, et les défenses cellulaires (monocytes, macrophages, polynucléaires neutrophiles et les cellules "natural killers") qui participent à la réaction inflammatoire.
Barrière naturelle
La limite extérieure du corps constitue une barrière efficace contre la plupart des agents microbiens qui ne peuvent, notamment, traverser la peau à l'état normal. On connaît l'importance de cette barrière : son absence se fait sérieusement sentir chez les grands brûlés, chez qui la prévention de l'infection par voie cutanée est une préoccupation majeure. La plupart des germes pénètrent par l'épithélium du nasopharynx, l'intestin, les poumons ou les voies génito-urinaires. Ces régions sont normalement protégées par divers mécanismes de défense. Par exemple, il existe dans différentes sécrétions une enzyme, appelée lysosome, capable de rompre certaines liaisons chimiques de la paroi de nombreuses bactéries et donc de les détruire.
Cellules phagocytaires
Lorsqu'un micro-organisme franchit un revêtement épithélial, il se trouve en présence des cellules phagocytaires du système réticulo-histiocytaire. Ces cellules, qui peuvent être de différents types, proviennent toutes des cellules souches de la moelle osseuse. Leur fonction est de capter toutes sortes de particules y compris les germes pathogènes, les phagocyter et enfin les détruire. Elles se trouvent placées en différents points stratégiques, sur le passage de ces particules. Les cellules de Kupffer sont situées le long des capillaires sinusoïdes du foie ou des cellules synoviales. Dans le sang, les cellules phagocytaires sont les polynucléaires neutrophiles et les monocytes. Les monocytes sont capables, en réponse à des signaux bien définis, de franchir les parois vasculaires pour aller migrer dans les tissus avoisinants où ils deviennent des macrophages tissulaires.
Cellules NK
Les cellules dites "Natural Killer" (NK) sont des leucocytes capables de reconnaître les altérations membranaires des cellules infectées par un virus et de lyser ces cellules. Les cellules NK sont activées par les interférons. Les interférons sont produits par des cellules parasitées par un virus. Ils sont capable de rendre résistantes aux virus des cellules non encore infectées.
Au cours d'une infection, la concentration d'un grand nombre de protéines plasmatiques augmente rapidement. Un exemple illustre bien ce phénomène, c'est la protéine C-réactive. En se fixant sur les bactéries, la protéine C-réactive prépare l'action du complément, qui a son tour facilite la captation des bactéries par les cellules phagocytaires (opsonisation). Le Complément est un ensemble d'une vingtaine de protéines plasmatiques interagissant entre elles ainsi qu'avec d'autres effecteurs de l'immunité.
Phénomène d'inflammation
L'inflammation est la réaction de l'organisme face à une agression comme celle que constitue une invasion microbienne. Elle se caractérise par 3 phénomènes majeurs :
- Un appel de sang vers la région infectée afin d'apporter de l'oxygène et du glucose
- Une augmentation de la perméabilité des capillaires par suite de la contraction des cellules endothéliales. Ceci autorise les grosses molécules à franchir plus facilement l'endothélium vasculaire, permettant aux médiateurs solubles de l'immunité d'atteindre le site infectieux.
- Une migration des leucocytes, en particulier les polynucléaires neutrophiles et dans une moindre mesure les macrophages, hors des vaisseaux capillaires jusqu'au site de l'infection.
Immunité spécifique
La situation se complique quand les cellules phagocytaires sont incapables de reconnaître l'agent infectieux, soit parce qu'elles ne possèdent pas le récepteur approprié, soit parce que le micro-organisme n'active pas le complément et ne peut, de ce fait, se fixer sur la cellule phagocytaire au moyen du recepteur C3b. Dans ces conditions, l'idéal pour l'organisme est de disposer d'un médiateur suffisamment flexible pour pouvoir s'ajuster par l'une de ses extrémités à l'agent infectieux et par l'autre à la cellule phagocytaire. Pour répondre à cette nécessité, sont apparues au cours de l'évolution, des molécules appelées anticorps.
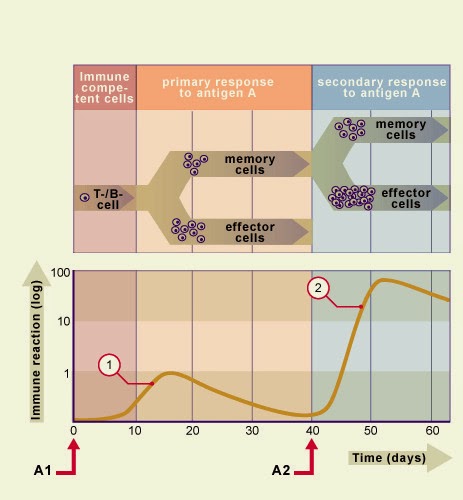 Les anticorps sont produits par les lymphocytes B. Un anticorps donné se fixe à un seul agent infectieux et peut se lier, par son autre extrémité, à des récepteurs de membrane de la cellule phagocytaire, les récepteurs Fc. On trouve les récepteur Fc sur les macrophages, les polynucléaires neutrophiles et sur toutes les cellules du système réticulo-histocytaire. Les anticorps étant capables d'activer le complément par la voie dite "classique", les agents infectieux se trouvent fréquemment recouverts à la fois d'anticorps et de C3b. Dans ce cas, la cellule phagocytaire reconnaît l'agent infectieux à la fois par l'intermédiaire de ses récepteurs Fc et de recepteur du C3b.
Les anticorps sont produits par les lymphocytes B. Un anticorps donné se fixe à un seul agent infectieux et peut se lier, par son autre extrémité, à des récepteurs de membrane de la cellule phagocytaire, les récepteurs Fc. On trouve les récepteur Fc sur les macrophages, les polynucléaires neutrophiles et sur toutes les cellules du système réticulo-histocytaire. Les anticorps étant capables d'activer le complément par la voie dite "classique", les agents infectieux se trouvent fréquemment recouverts à la fois d'anticorps et de C3b. Dans ce cas, la cellule phagocytaire reconnaît l'agent infectieux à la fois par l'intermédiaire de ses récepteurs Fc et de recepteur du C3b.